Видео:Влияние внешних и внутренних факторов на транспирацию. 9 класс.Скачать

Площадь испаряющей поверхности листа
Низкая влажность воздуха около листа способствует транспирации, поскольку диффузионный градиент водяного пара (градиент водного потенциала) между межклетниками и окружающей атмосферой становится круче. С повышением концентрации водяного пара в воздухе, т. е. с увеличением влажности, диффузионный градиент становится более пологим. Водный потенциал атмосферы убывает также с увеличением высот над уровнем моря; это связано с понижением атмосферного давления. Поэтому у высокогорных растений часто наблюдаются ксероморфные признаки, снижающие интенсивность транспирации.
К таким признакам относятся, например, погруженные устьица, т. е. устьица находящиеся на дне борозд или складок эпидермиса, в которых локально создается повышенная влажность, что замедляет транспирационные потери воды. У некоторых видов, например у Ammophila (песколюба), вся листовая пластинка скручивается в трубку, так что устьица оказываются внутри этой трубки, где возникает влажная атмосфера. Если лист покрыт густо расположенными эпидермальными волосками или чешуйками, то между ними задерживается диффундирующий из устьиц пар, и скорость транспирации также снижается.
Движение воздуха
В неподвижном воздухе вокруг листа формируется слой насыщенного влагой воздуха, и диффузионный градиент между межклетниками и окружающей атмосферой становится относительно пологим. При малейшем движении воздуха этот слой сдувается, поэтому интенсивность транспирации при ветреной погоде повышается, причем усиление транспирации заметнее всего, когда ветер слабый. При сильном ветре устьица закрываются, и транспирация прекращается.

Как уже указывалось, опущение листа или чешуйки на его поверхности создают зону спокойного воздуха и таким образом несколько замедляют транспирацию.
Освещенность влияет на транспирацию, поскольку устьица обычно открыты на свету и закрыты в темноте. Таким образом, ночью растения теряют сравнительно мало воды (через кутикулу или чечевички). Утром устьица раскрываются, и интенсивность транспирации возрастает.
Мы уже говорили о том, как влияют на интенсивность транспирации некоторые ксероморфные адаптации. Ниже приводится еще несколько примеров такого влияния эндогенных, т. е. внутренних, несредовых, факторов.
Площадь испаряющей поверхности и отношение этой поверхности к объему растения
Транспирация усиливается при возрастании общей площади поверхности листьев растения и отношения этой площади к его объему. Поверхность листьев уменьшается с приобретением ими чешуе- или игловидной формы, как у хвойных, или при редукции их до колючек, как у кактусов. К этому же ведет и мелколистность, свойственная ксерофитам. Ксероморфной адаптацией является также сбрасывание листвы в сухой или холодный сезон. В последнем случае это важно, поскольку вода в почве замерзает и становится недоступной растению.
Отношение испаряющей поверхности к объему растения уменьшается, когда его главной фотосинтезирующей частью становится стебель, как у кактусов. На рисунке показаны примеры характерного уменьшения листовой поверхности у кактусов и других суккулентов.
Видео:Что такое транспирация у растений? Испарение воды листьями.Скачать

Транспирация

У растений, кроме нижнего двигателя водного тока (корневое давление), есть еще верхний концевой двигатель — испарение воды листьями. Вследствие этого происходит поступление и передвижение воды по растению. Испарение воды растением называется транспирацией. Ее можно рассматривать как физический процесс — переход воды в парообразное состояние и диффузию образовавшегося пара в окружающее пространство.
Испарение воды, растением можно значительно уменьшить, если выращивать его в атмосфере, насыщенной водяным паром, например в условиях оранжереи или теплицы. Снижению транспирации способствуют также освежающие поливы дождеванием в утреннее время, когда происходит насыщение воздуха вокруг растения парообразной влагой.
Растения имеют ряд приспособлений и анатомических особенностей — кутикулу, восковой налет, покровные волоски, которые уменьшают транспирацию. Значительно сокращает ее опушенность нижней поверхности листьев. Однако прекращение транспирации отрицательно сказывается на фотосинтезе, поскольку при диффузии и испарении воды в окружающую атмосферу через устьица в растение поступает углекислый газ.
Между водным режимом и углеродным питанием, т. е. между поступлением углекислого газа в растение и транспирацией, существует глубокое внутреннее противоречие. С одной стороны, интенсивная транспирация может привести к отрицательным явлениям, а с другой — способствует большему притоку СО2, лучшему углеродному питанию растений.
Транспирация создает автоматичность водного тока: поступление воды в растение и испарение ее. Без транспирации растение не будет обеспечено водой, поскольку корневое давление подает незначительное количество ее. Транспирация способствует передвижению минеральных веществ в растении, которые поглощаются из почвы корнями. Кроме того, благодаря ей снижается температура растения. Без транспирации растения перегревались бы и различные физиологические и биохимические процессы не могли бы нормально происходить.
Для естественного функционирования растений, особенно в период их цветения и плодоношения, необходима достаточная насыщенность клеток водой. В определенной мере это обеспечивается транспирацией. Следовательно, это важный и необходимый физиологический процесс.
Транспирация характеризуется следующими показателями. Количество воды, испаряемой растением с единицы листовой поверхности в единицу времени, называется интенсивностью транспирации. Она может быть выражена в граммах на 1 м 2 или 1 см 2 за один час. Испаряемую воду можно отнести к массе листьев. Это также будет показателем интенсивности транспирации. Обычно для большинства растений она в среднем составляет 15-250 г на 1 м 2 днем и 1-20 г на 1 м 2 ночью. Интенсивность транспирации прямо пропорциональна разности между концентрацией водяного пара у испаряющей поверхности и содержанием его в атмосфере (г/см 3 ), площади испаряющей поверхности и обратно пропорциональна сумме диффузионных сопротивлений: $$large Тр = frac$$
где Тр — интенсивность транспирации; ΔС — градиент концентрации водяного пара между транспирирующей поверхностью и окружающим воздухом; Σr — сумма диффузионных сопротивлений листа — пограничного слоя, устьичного и кутикулярного.
Сопротивление пограничного слоя (rа) у мелких листьев при отсутствии ветра достигает 1 с/см, у крупных — 3 с/см; при скорости ветра около 2 м/с сопротивление составляет меньше 0,3 с/см. Устьичное диффузионное сопротивление (rs) зависит от степени открытия устьиц. У травянистых сельскохозяйственных растений оно находится в пределах 0,8-2 с/см. У листопадных деревьев, кустарников и большинства злаков минимальное диффузионное сопротивление листьев составляет 2-4 с/см, у тенелюбивых трав, степных злаков и вечнозеленых лиственных деревьев — 3-5, у хвойных и суккулентов — 6-12 с/см.
Кутикулярное сопротивление диффузии у большинства растений обычно равно 20-100 с/см, а у хвои с усиленной защитой от транспирации оно может достигать 400 с/см (В. Лархер).
Транспирация является физиологически регулируемым процессом. Изменением ширины устьичных отверстий растение регулирует водообмен и одновременно контролирует поступление СО2 в лист. Устьичная газопроводимость листа (l/rs — величина, обратная устьичному сопротивлению) прямо пропорциональна площади устьичных щелей и возрастает с увеличением их ширины.
Скорость использования водного запаса характеризуется количеством затраченной растением воды в единицу времени и выражается в процентах к общему ее количеству в растении. Расход запаса воды в растении колеблется от 10 до 80%. Количество созданного сухого вещества на 1 кг транспирированной воды служит показателем продуктивности транспирации. Она характеризуется образованием 1-8 г (в среднем 3 г) сухого вещества при прохождении 1 кг воды.
Транспирационный коэффициент показывает, сколько воды растение затрачивает на построение единицы сухого вещества. Это величина, обратная продуктивности транспирации. Транспирационный, коэффициент у растений находится в пределах 125-1000, а средний равен 300, т. е. на 1т урожая затрачивается 300 т воды.
Условия выращивания растений оказывают большое влияние на величину транспирационного коэффициента. Исследования показали, что на неудобренной почве растение расходует воду менее продуктивно, чем на удобренной. В последнем случае количество воды, необходимое для создания единицы сухого вещества растением пшеницы, уменьшилось почти в 2 раза. При большом содержании воды в растении и удобрении создается и большая масса урожая. Следовательно, при обеспечении растений питательными веществами они более продуктивно расходуют воду. При прямом солнечном свете на создание единицы сухой массы растение затрачивает наименьшее количество воды, а при рассеянном — намного больше, что объясняется слабой интенсивностью фотосинтеза.
Устьичная транспирация. Исследования анатомического строения листьев и других частей растения показали, что вода испаряется главным образом через устьица. Устьичная транспирация состоит из двух фаз: собственно испарения воды с поверхности влажных клеток мезофилла и диффузии через устьица водяного пара, образовавшегося в межклетниках. Процесс испарения воды осуществляется благодаря тому, что молекулы воды при наличии градиентов способны преодолевать силы натяжения и силы внутреннего сцепления частиц жидкости и, отрываясь от нее, переходить в виде пара в воздух.
Наблюдая этот процесс в сосудах разного диаметра, можно увидеть, что диффузия молекул в парообразном состоянии при этих условиях имеет особенности. Испарение воды возле краев, сосуда малого диаметра более интенсивное, чем в средней части, потому что молекулы водяного пара здесь двигаются не только перпендикулярно, но и под некоторым углом — веерообразно, трение одних частиц воды о другие меньше, следовательно, испарение будет происходить быстрее. Оказывается, что вследствие этого меньшие площади испаряют воду сравнительно быстрее, чем большие.
И. Стефан и Б. И. Срезневский установили закономерность, согласно которой испарение из малых отверстий происходит пропорционально периметру, и предложили формулу $$large upsilon = K4a; frac
,$$
где ʊ — количество воды, испаряемой с единицы площади; К — коэффициент диффузии исследуемой жидкости; а — радиус испаряющей поверхности; F — упругость пара, который насыщает воздух при температуре испаряющей поверхности; f — фактическая упругость пара в воздухе; Р — атмосферное давление.
Согласно этой формуле, из двух одинаковых по форме сосудов, например плоских чашек, диаметр которых различается в 2 раза, а площадь в 4 раза, большая чашка будет испарять лишь в 2 раза больше воды, чем меньшая.
Устьица составляют не более 1% всей площади листа. На 1 мм 2 листовой поверхности их насчитывается от 50 до 500 и более, в зависимости от вида и сорта растения и стороны листа. Каждое устьице можно представить в виде очень маленького сосудика. Поэтому несколько небольших отверстий в какой-либо перегородке будут пропускать пары воды быстрее, чем одно большое отверстие, равное им по диаметру. Таким образом, чем больше отношение периметра к площади (а оно тем больше, чем меньше диаметр), тем быстрее происходят испарение и диффузия. В этом случае большое значение имеет так называемый краевой эффект (рис. 16).
Если взять два сосуда одинакового диаметра, один оставить открытым, а другой покрыть перегородкой с мелкими отверстиями, то испарение и диффузия будут осуществляться более интенсивно во втором случае, поскольку величина испарения здесь пропорциональна диаметру мелких отверстий. Поэтому транспирация с поверхности листа в количественном выражении приближается к испарению воды с открытой поверхности такой же площади. Сравнительное определение величины транспирации — так называемая относительная транспирация — у некоторых растений показало, что она составляет 0,9, а средняя — 0,4-0,5, т. е. транспирация листа соответствует испарению с половины открытой водной поверхности, равной площади листа.

Рис. 16.Схема, объясняющая краевой эффект:
маленькие отверстия (вверху), например устьичная щель, имеют большее диффузионное поле, чем равная площадь открытой поверхности (по Э. Либберту).
Устьичная регуляция транспирации. У большинства растений устьица замыкаются клетками полулунной или бобовидной формы. У растений семейства Мятликовые (Злаковые) устьица замыкаются не бобовидными клетками, а продольными (рис. 17).
Устьица могут закрываться и открываться. Их движение обусловлено различными факторами. Необходимо указать на некоторые морфологические особенности строения клеток, замыкающих устьица. Оболочки замыкающих клеток, примыкающие к устьичным щелям, плотнее и толще. Основным фактором, обусловливающим движение устьиц, является содержание воды в листе. При достаточном ее количестве в растении устьица открыты. На свету они у большинства растений тоже открыты, а в темноте закрываются.
В клетках, замыкающих устьица, на свету крахмал превращается в сахар, а в темноте, наоборот, сахар переходит в крахмал. Сахар как осмотически действующее вещество способствует поглощению воды клетками, замыкающими устьица, и последние открываются. В темноте накапливается крахмал, поэтому поглощения воды не происходит и устьица закрываются. Движение устьиц регулируется комплексом факторов (температура, осмотическое давление, интенсивность освещения и качество света — длинноволновые и коротковолновые лучи, увлажнение и др.). При этом в зависимости от условий и вида растения сочетание внешних и внутренних факторов может быть разным.
Высказывается также предположение, что снижение обводненности листьев приводит в клетках, замыкающих устьица, к накоплению абсцизовой кислоты С15Н20О4, которая ингибирует синтез α-амилазы. Гидролиз крахмала прекращается, вследствие чего снижается осмотическое давление и устьица закрываются.

Рис. 17. Устьица в открытом (вверху) и закрытом (внизу) состоянии:
А — двудольного растения; Б — злака.
Гистохимические исследования показали, что при уменьшении водного потенциала в замыкающих клетках устьиц листьев подсолнечника при сниженной водообеспеченности в условиях освещения пропорционально уменьшалось и содержание в них К + , возрастало сопротивление листьев воздуху, что указывало на закрывание устьиц. После дегидратации листьев устьица открывались только через несколько часов, когда К + вновь поступал в клетки. Тургесцентные сегменты листьев, плавающие в растворе с добавлением абсцизовой кислоты (0,1 мМ), теряли К + из замыкающих клеток, и устьица быстро и плотно закрывались, как и при подсушивании.
Следовательно, в условиях освещения при низком водном потенциале открытие устьиц регулируется не только потерей воды тканями, но и потерей К + в замыкающих клетках. Это, в свою очередь, уменьшает тургорный градиент между замыкающими и окружающими клетками, в результате чего устьица закрываются.
Таким образом, установлено влияние калия на степень раскрытия устьиц. Обнаружена положительная корреляция между концентрацией калия в замыкающих клетках устьиц и величинной устьичного отверстия. Считается, что анионами, обеспечивающими баланс калия в замыкающих клетках, являются малат и хлор.
Интересно отметить, что только при содержании калия в сухом веществе более 1% транспирационный коэффициент низкий, а эффективность использования воды высокая.
Снабжение растений достаточным количеством калия позволяет им противостоять водному стрессу благодаря механизму саморегуляции и уменьшению транспирации.
Движение устьиц связано также с изменением вязкости протопласта замыкающих клеток. Об этом свидетельствуют различные формы плазмолиза в них. При открытых устьицах плазмолиз имеет выпуклую форму, а при закрытых — судорожную.
Различают также устьичные движения: фотоактивную реакцию, если устьица на свету открываются, а в темноте закрываются; гидроактивную реакцию, когда устьица закрываются при сильной потере воды листом; гидропассивную реакцию, если, например, в дождливую погоду при насыщении клеток листа водой устьица закрываются. Это происходит при значительном увеличении объема эпидермиса клеток, сдавливающих устьичные клетки.
Общая поверхность межклетников в листе очень большая. У теневыносливых растений внутренняя поверхность межклетников превышает внешнюю в 7-10 раз, у гидрофитных растений в 12-19, ксерофитных — в 17-30 раз. Благодаря большей испаряющей поверхности в межклетниках отмечается интенсивный выход водяного пара через устьица наружу.
У кукурузы на 1 см 2 нижнего эпидермиса листа насчитывается 7684 устьица, верхнего — 9300. Средняя площадь одного устьичного отверстия равна 89 мкм 2 . На одном растении кукурузы насчитывается около 104 100 000 устьиц. Такое большое их количество и обеспечивает интенсивную диффузию водяного пара и транспирацию.
Периодичность устьичных движений и кутикулярная транспирация. Исследованиями устьичных движений у разных растений установлено, что в ясную, но не очень сухую погоду у большинства растений устьица открываются на рассвете и устьичная щель достигает максимума в утренние часы, в полдень она начинает немного сужаться и закрывается во время захода солнца (рис. 18, 19). В пасмурную погоду устьица обычно открыты, но не так широко, как в ясную. В сухую же погоду они, как правило, открывшись утром, в 10-11 ч закрываются.

Рис. 18. Устьица яблони:
А — в закрытом состоянии; Б — в открытом состоянии; В — поперечный разрез.
У разных растений устьица функционируют неодинаково. Так, у картофеля, капусты, кормовой свеклы преобладает открытое состояние устьиц, и при достаточной водообеспеченности они не закрываются ни днем, ни ночью. У хлебных злаков они закрываются вечером и на протяжении ночи не открываются. У растений, которые имеют устьица на обеих сторонах листа, на верхней стороне они закрываются позже и на более короткий срок, возле верхушки листа устьица открываются раньше и закрываются позже, чем возле его основания.

Рис. 19. Часть нижнего эпидермиса листа герани:
1 — устьичная щель; 2 — замыкающая клетка.
Исследования В. Г. Карманова и С. Н. Мелещенко показали, что устьица могут быть более открытыми в тот момент, когда водоснабжение листа уменьшается, и менее открытыми, когда лист находится в достаточно тургесцентном состоянии. Это парадоксальное явление объясняется большей проницаемостью и скоростью водных токов по микрокапиллярам клеточных целлюлозных оболочек, чем через цитоплазму и вакуоли (трансвакуолярно).
В результате переноса микроскопических количеств воды по клеточным оболочкам возрастает содержание ее в клетках, замыкающих устьица, апертура устьичных щелей увеличивается и транспирационные расходы воды также могут возрастать. Течения, которые создаются нижним двигателем водного тока, незначительны, и идут они главным образом по клеточным оболочкам, составляющим 10% объема ткани листа. Это пример механизма прямой связи между нижним и верхним двигателями водного тока в растении. Но существует и обратная связь: верхний двигатель → нижний двигатель водного тока, а именно апертура устьичных щелей — фотосинтез — отток ассимилятов в корни — корневое давление.
Внеустьичная транспирация. Кроме устьичной, существует еще так называемая внеустьичная транспирация. Теряя значительное количество воды в процессе транспирации, растение обычно регулирует отдачу воды в соответствии с ее поступлением через корневую систему. Поэтому создание бездефицитного водного баланса, т. е. пропорциональности между расходом воды и ее поступлением, — одно из необходимых условий существования всех растений, особенно в условиях засухи.
Следует отметить, что уменьшение диаметра устьичной щели на 50-75% максимальной ее величины еще не влияет заметно на транспирацию.
Таким образом, роль устьичных движений в регулировании транспирации довольно незначительна. Лист покрыт сплошным слоем эпидермальных клеток, внешние стенки которых имеют кутикулу и малопроницаемы для воды и водяного пара. Если нижнюю сторону листа древесного растения, у которого здесь обычно расположены устьица, покрыть вазелином, то незначительная транспирация все же будет наблюдаться. Это и есть кутикулярная транспирация, она в 10-20 раз слабее устьичной.
У теневыносливых растений кутикулярная транспирация достигает значительных размеров — до 50%.
Внеустьичная регуляция транспирации у растений заключается в задержке испарения или уменьшении содержания воды в оболочках клеток, выстилающих межклетники. Таким образом, недостаток воды в листьях вследствие большой транспирации или недостаточного поступления воды из почвы является причиной сокращения транспирации, которая не зависит от сужения устьичных щелей. Такое явление получило название начинающегося подсыхания.
Водный баланс и завядание. Одним из наиболее динамичных процессов в растении является водный обмен, который находится в тесной корреляции с другими процессами его жизнедеятельности. Водный баланс — это поступление и расходование воды растением. При умеренной транспирации и достаточном поступлении воды в растение создается благоприятный водный баланс. В ясный солнечный день это равновесие нарушается и в растении возникает водный дефицит, который обычно составляет 5-10%. Такой дефицит считается вполне нормальным и не приносит особого вреда растению.
При интенсивной транспирации или иссушении почвы, когда поступление воды в растение прекращается, происходит значительная потеря ее растительными клетками, которая не пополняется поглощением влаги из почвы, в результате чего создается водный дефицит, часто наблюдаемый у растений в наиболее жаркие часы суток. При водном дефиците листья теряют тургор, завядают, повисают.
Водный дефицит листьев можно определить по формуле: $$large W = (1 — frac)100,$$
где W —водный дефицит, %; m — масса высечек до насыщения их водой; m1 — масса высечек после насыщения их водой; экспозиция насыщения 60 мин.
У некоторых растений, имеющих в органах большое количество механических тканей, например у бессмертника (род Helichrysum), внешний вид при водном дефиците, значительной потере воды и даже при гибели не изменяется.
Наблюдения показали, что обычно на рассвете внутренний градиент в растении и почве почти выравнивается, уравновешиваются водные потенциалы растения и почвы. В утренние часы, когда листья начинают транспирировать, водный потенциал становится несколько меньшим, чем на рассвете» однако поступление воды в растение начинается, когда создается необходимый градиент водных потенциалов от листьев к поверхности раздела корень — почва.
Завядание бывает временным и длительным.
Временное завядай не отмечается обычно при сильной жаре и сухости воздуха. Часто в полдень можно наблюдать потерю тургесцентного состояния листьями многих растений. При временном завядании фотосинтез и рост приостанавливаются. Оно не приносит большого вреда: на протяжении ночи растение восстанавливает нормальное состояние.
Длительное завядание наступает, когда почва не содержит доступной для растений влаги. При этом тургесцентное состояние листьев не восстанавливается, создается остаточный водный дефицит, корневые волоски отмирают, вследствие чего даже после полива растений поглощение воды происходит очень медленно, и лишь с появлением новых корневых волосков водоснабжение приходит в норму. При длительном завядании обезвоживаются эмбриональные клетки тканей, что приводит к глубокому нарушению свойств протопласта и способности клеток к росту. Ферменты, регулирующие превращение крахмала в сахар, подвергаются изменениям необратимого характера. При длительном завядании протопласта изменяются свойства коллоидов: увеличивается проницаемость протопласта, и при погружении растения в воду наблюдается, значительный экзоосмос электролитов и органических «веществ». Плазмолиз в клетках таких тканей проходит быстро, уменьшается дисперсность биоколлоидов. Кроме того, повреждаются зеленые пластиды, снижается их ассимиляционная способность, приостанавливается фотосинтезирующая активность растений, усиливается дыхание растительных тканей, а в период плодоношения задерживается налив зерна. Вода растущих частей растения оттягивается клетками к органам, имеющим более высокую концентрацию осмотически активных веществ. Именно этим и объясняется так называемый «захват» зерна, при котором вода из колоса перемещается в ткани листьев и стебля. Если растение находится некоторое время в завядшем состоянии в фазе молочной или восковой спелости, то зерно хлебных злаков становится щуплым, а у хлопчатника происходит сбрасывание бутонов и даже завязей.
У светолюбивых растений, например подсолнечника, кукурузы, может утрачиваться 25-30% имеющейся в них воды без внешних признаков завядания: клеточные оболочки у таких растений обладают способностью сильно растягиваться, что не отражается на их внешнем состоянии. У растений, произрастающих в тени, например под пологом леса, при потере 2-3% воды уже происходит завядание, так как их клеточные оболочки неспособны сильно растягиваться.
Длительное нарушение водного баланса у растений приводит к изменениям некоторых физиологических процессов. Завядание растений вследствие недостатка влаги вызывает нарушение нормального обмена веществ, изменяются осмотические свойства, значительно увеличивается проницаемость цитоплазмы, возрастают сосущая сила, интенсивность дыхания, задерживаются процессы роста, уменьшаются листовая поверхность и ассимиляционная способность организма — все это приводит к резкому снижению урожайности.
Поэтому относительное содержание воды в растении, определяемое главным образом разностью между количеством влаги, расходуемой на транспирацию, и количеством влаги, поглощаемой корневой системой, что лежит в основе концепции водного баланса, является важным физиологическим показателем, при описании состояния воды в растении и уровня стресса при засухе.
Зимняя транспирация. Травянистые многолетние растения почти не расходуют воду зимой, поскольку их органы (корни, клубни, корневища) защищены почвой и снежным покровом. Древесные породы обладают способностью противодействовать расходу воды, транспирации не только летом, но и зимой. Если принять за единицу расход воды годичным побегом лиственницы (Larix sibirica), то показатели транспирации древесных пород, выросших в одинаковых условиях, могут быть расположены в виде ряда увеличивающихся цифр. Чем дальше на север, тем больше древесные породы приближаются к лиственнице: зимой она транспирирует слабее.
У древесных пород имеется пробковый слой, который обрадуется обычно при дозревании побега. Он защищает растение от транспирации, а не от мороза, как обычно считают. В этом можно легко убедиться, если на одном побеге бузины срезать кору вблизи от почек, а на втором также снять коровую паренхиму и место среза покрыть станиолью. Зимой через некоторое время почки, расположенные близко к срезу, погибнут, а на побеге, где срез был защищен станиолыо, останутся живыми. Этот опыт наглядно иллюстрирует значение пробкового слоя у древесных растений в зимний период. Иглы хвойных пород покрыты толстым слоем кутикулы с налетом воска, в зимний период устьица в них всегда закрыты.
В клетках зимующих побегов происходят изменения, которые повышают сопротивление отдаче воды: уменьшается проницаемость протопласта, а также увеличивается количество связанной коллоидами воды.
Определение влажности игл и побегов хвойных пород (опыты Л. А. Иванова под Ленинградом) показало, что потеря влаги в зимний период составляет не более 3-5%, а у южных пород — 10-15%. Гибель от высыхания наблюдается только при потере около 50% влаги, которая содержалась в побеге. Высыхание в зимний период наблюдается главным образом у сеянцев. Это явление отмечается в восточной части СССР, где снежный покров небольшой и число солнечных дней значнтельное. Исследованиями Л. А. Иванова установлено, что в древесине пня после срезания кроны наблюдается повышение влажности. Причиной этого может быть только корневое давление. В том случае, когда были перерублены корни, увеличение влажности в тканях пня приостанавливалось. Установлено также, что влажность древесины у основания ствола всегда минимальная и возрастает возле кроны. Причиной этого является увеличение свободного пространства (пустот) в клетках древесины, а следовательно, и ее влагоемкости.
Зимняя транспирация происходит также у озимых хлебных злаков и многолетних трав.
Видео:Лист. Обнаружение испарения воды листьямиСкачать

Проект по биологии на тему «ВЛИЯНИЕ ПЛОЩАДИ ПОВЕРХНОСТИ ЛИСТЬЕВ РАЗНЫХ ВИДОВ РАСТЕНИЙ НА ТРАНСПИРАЦИЮ» (9 класс)
Обращаем Ваше внимание, что в соответствии с Федеральным законом N 273-ФЗ «Об образовании в Российской Федерации» в организациях, осуществляющих образовательную деятельность, организовывается обучение и воспитание обучающихся с ОВЗ как совместно с другими обучающимися, так и в отдельных классах или группах.
Развитие управляющих функций мозга ребёнка: полезные советы и упражнения для педагогов
Сертификат и скидка на обучение каждому участнику
Муниципальное автономное общеобразовательное учреждение
Школьная научно-практическая конференция
ВЛИЯНИЕ ПЛОЩАДИ ПОВЕРХНОСТИ ЛИСТЬЕВ РАЗНЫХ ВИДОВ РАСТЕНИЙ НА ТРАНСПИРАЦИЮ
Выполнила: Еремеева Кристина
ученица 9б класса
Руководитель: Угрюмова Екатерина Анатольевна
учитель высшей категории
I Транспирация и ее особенности
Понятие транспирации и ее виды………………………………………5-6
Условия, влияющие на испарение воды ………………………………6-9
II Роль листьев в испарении воды растениями
2.1 Лист как орган транспирации ………………………………………….10-12
2.2 Биологическое значение транспирации ……………………………….13-14
III Практическая работа «Определение общей площади поверхности
листьев дуба, березы и тополя» …………………………………………….14
В настоящее время наблюдается процесс опадания листьев в июле, августе, несмотря на то, что в летние месяцы такое явления как листопад не характерно.
В связи с тем, что лето в Нижнем Тагиле бывает очень жаркое, а именно температура высокая, много света, низкая влажность почвы – это приводит к снижению транспирации. Недостаток испарения нарушает правильное распределение питательных веществ и препятствует движению воды и питательных веществ от корней к побегам.
Транспирация спасает растение от перегрева, который ему грозит на прямом солнечном свете. Температура сильно транспирирующего листа может примерно на 7°С быть ниже температуры листа завядающего, нетранспирирующего. Это особенно важно в связи с тем, что перегрев, разрушая хлоропласты, резко снижает процесс фотосинтеза (оптимальная температура для процесса фотосинтеза около 30-33°С). Именно благодаря высокой транспирирующей способности многие растения хорошо переносят повышенную температуру. Большая поверхность листьев имеет огромное значение для питания растений и для испарения большого количества воды.
Чем больше развита листовая поверхность, тем больше общая потеря воды. Поверхность листа покрыта порами, называемыми устьицами и у большинства растений большая часть устьиц находится на нижней части листа. Устьица ограничены замыкающими клетками и сопровождающими клетками (вместе известными как устьичный комплекс), которые открывают и закрывают поры. Транспирация проходит через устьичные щели и может рассматриваться как необходимая «цена», связанная с открытием устьиц для доступа углекислого газа, необходимого для фотосинтеза. Транспирация также охлаждает растение, изменяет осмотическое давление в клетках и обеспечивает движение воды и питательных веществ от корней к побегам.
Однако в процессе естественного отбора у растений одновременно с большей листовой поверхностью выработалась компенсирующая способность к меньшему испарению с единицы поверхности листа (меньшая интенсивность транспирации). У кактусов листья превращены в колючки, которые защищают их от поедания животными и испаряют мало воды.
Листья многих растений сухих мест имеют густое опушение, восковой налет и испаряют мало влаги.
Таким образом, была сформулирована тема исследовательской работы
Влияние площади поверхности листьев разных видов растений на транспирацию
Объект: особенности транспирации
Предмет: лист как орган транспирации
Цель: Определение роли транспирации в жизни растений, ее зависимость от площади поверхности листьев разных видов растений (дуб, береза, тополь)
1.Изучить литературу по проблеме исследования.
2. Изучить понятие транспирации и охарактеризовать ее виды
3. Выявить условия, влияющие на испарение воды
4. Рассмотреть значение листа в процессе транспирации
5. Сформулировать биологическое значение транспирации
6. Сравнить площадь испаряющей поверхности листьев разных видов растений (дуб, береза, тополь)
Гипотеза исследовательской работы: от площади поверхности листа зависит рост растений в разных температурных условиях.
Методы исследования: наблюдение, анализ, синтез изученного материала.
Практическая часть данной работы : проведение лабораторной работы на тему «Определение общей площади поверхности листьев дуба, березы и тополя».
I Транспирация и ее особенности
1.1 Понятие транспирации и его виды
Транспирация — процесс движения воды через растение и её испарение через наружные органы растения, такие как листья, стебли и цветы. Вода необходима для жизнедеятельности растения, но только небольшая часть воды, поступающей через корни используется непосредственно для нужд роста и метаболизма. Оставшиеся 99-99,5 % теряются через транспирацию. Поверхность листа покрыта порами, называемыми устьицами и у большинства растений большая часть устьиц находится на нижней части листа. Устьица ограничены замыкающими клетками и сопровождающими клетками (вместе известными как устьичный комплекс), которые открывают и закрывают поры. Транспирация проходит через устьичные щели и может рассматриваться как необходимая «цена», связанная с открытием устьиц для доступа углекислого газа, необходимого для фотосинтеза. Транспирация также охлаждает растение, изменяет осмотическое давление в клетках и обеспечивает движение воды и питательных веществ от корней к побегам.
Вода поглощается корнями из почвы с помощью осмоса и движется в ксилеме наверх вместе с растворенными в ней питательными веществами. Движение воды от корней к листьям частично обеспечивается капиллярным эффектом, но в основном происходит за счет разности давлений. В высоких растениях и деревьях, гравитация может быть преодолена только за счет уменьшения гидростатического давления в верхних частях растения из-за диффузии воды через устьица в атмосферу. [Приложение №1]
Охлаждение достигается путём испарения с поверхности растения воды, у которой высокая удельная теплота парообразования [Геннель 2010-470].
Выделяют следующие виды транспирации:
— кутикулярная транспирация. Снаружи листья имеют однослойный эпидермис, внешние стенки клеток которого покрыты кутикулой и воском, образующие эффективный барьер на пути движения воды. На поверхности листьев часто развиты волоски, которые также влияют на водный режим листа, так как снижают скорость движения воздуха над его поверхностью и рассеивают свет и тем самым уменьшают потери воды за счет транспирации.Интенсивность кутикулярной транспирации варьирует у разных видов растений. У молодых листьев с тонкой кутикулой она может составлять около половины всей транспирации. У зрелых листьев с более мощной кутикулой кутикулярная транспирация равна 1/10 общей транспирации. В стареющих листьях из-за повреждения кутикулы она может возрастать. Таким образом, кутикулярная транспирация регулируется главным образом толщиной и целостностью кутикулы и других защитных покровных слоев на поверхности листьев. Кутикулярная транспирация обычно составляет около 10% от общей потери воды листом.
-устьичная транспирация. Основная часть воды испаряется через устьица. Устьица играют важную роль в газообмене между листом и атмосферой, так как являются основным путем для водяного пара, углекислого газа и кислорода. Устьица находятся на обеих сторонах листа. Есть виды растений, у которых устьица располагаются только на нижней стороне листа. В среднем число устьиц колеблется от 50 до 500 на 1 мм². Транспирация через устьица идет почти с такой же скоростью, как и с поверхности чистой воды. Это объясняется законом И. Стефана: через малые отверстия скорость диффузии газов пропорциональна не площади отверстия, а диаметру или длине окружности. Поэтому, хотя площадь устьичных отверстий мала по отношению к площади всего листа (0,5-2 %), испарение воды через устьица идет очень интенсивно [Гэлстон 2009-380].
1.2 Условия, влияющие на испарения воды
Прежде всего испарение зависит от ненасыщенности атмосферы парами воды, или дефицита влажности. Это в целом справедливо и для транспирации. Однако надо учесть, что при недостатке воды в листе вступает в силу устьичная и внеустьичная регулировка, благодаря чему влияние внешних условий сказывается в смягченном виде и транспирация начинает возрастать медленнее. Несмотря на это, общая закономерность зависимости транспирации от насыщенности водой атмосферы остается справедливой. Чем меньше относительная влажность воздуха, тем выше интенсивность транспирации.
Следующим фактором среды, оказывающим влияние на процесс транспирации, является температура. С повышением температуры значительно увеличивается количество паров воды, которое насыщает данное пространство. Возрастание пространства приводит к повышению дефицита влажности. В связи с этим с повышением температуры транспирация увеличивается. Сильное влияние на транспирацию оказывает свет. Это связано с несколькими причинами: 1. На свету повышается температура листа и это вызывает усиление процесса транспирации. На транспирацию влияют поглощенные лучи, которые и вызывают повышение температуры. В связи с этим действие света на транспирацию проявляется тем сильнее, чем выше содержание хлорофилла. У зеленых растений даже рассеянный свет повышает транспирацию на 30 — 40%.
2. Под влиянием света увеличивается проницаемость цитоплазмы для воды, что также, естественно, увеличивает скорость ее испарения. 3. Под влиянием света устьица раскрываются. Все это вместе приводит к тому, что на свету транспирация идет во много раз интенсивнее, чем в темноте.
На интенсивность процесса транспирации оказывает влияние влажность почвы. С уменьшением влажности почвы транспирация уменьшается. Чем меньше воды в почве, тем меньше ее в растении. Уменьшение содержания воды в растительном организме автоматически снижает процесс транспирации, в силу устьичной и внеустьичной регулировки. В этой связи имеет значение и величина осмотического потенциала почвенного раствора. Чем она выше, тем ниже при прочих равных условиях интенсивность транспирации.
Ветер, перемешивая слои воздуха, очень сильно увеличивает скорость испарения. Ветер оказывает влияние и на транспирацию, правда по сравнению с испарением в несколько ослабленной форме. Поскольку обычно ветер не проникает внутрь листа, то под его влиянием возрастает в основном третий этап транспирации, т. е. перенос насыщенного водой воздуха от поверхности листа в более дальние слои атмосферы. В силу указанных причин при ветре усиливается прежде всего кутикулярная транспирация. Естественно поэтому, что более сильное действие ветер оказывает на транспирацию тех растений, где кутикула развита слабее. Сильнее, чем обычно, ветер сказывается на транспирации при суховеях. При суховеях ветер сгибает и разгибает листья и горячий воздух врывается в межклетники. Этим вызывается усиление транспирации уже на первом ее этапе.
Транспирация зависит как от внешних, так и от внутренних факторов, прежде всего от содержания воды в листьях. Всякое уменьшение содержания воды уменьшает транспирацию. Транспирация изменяется в зависимости от концентрации и осмотического давления клеточного сока. Молекулы воды удерживаются осмотическими силами. Чем концентрированнее клеточный сок, тем слабее транспирация. Интенсивность транспирации зависит от эластичности (способности к обратимому растяжению) клеточных стенок. Если клеточные стенки малоэластичны, то уже небольшая потеря воды приводит к сокращению объема клетки до минимума. В этот период клеточные оболочки не растянуты и не оказывают сопротивления, сосущая сила резко возрастает и становится равной всей величине осмотического давления. Возрастание сосущей силы клетки приводит к уменьшению транспирации [ Горышена 2011-345 ].
Транспирация изменяется в зависимости от величины листовой поверхности, а также при изменении соотношения корни/побеги.
Чем больше развита листовая поверхность, тем больше общая потеря воды. Однако в процессе естественного отбора у растений одновременно с большей листовой поверхностью выработалась компенсирующая способность к меньшему испарению с единицы поверхности листа (меньшая интенсивность транспирации).
С увеличением возраста интенсивность транспирации, как правило, падает. Смена дня и ночи, смена условий в течение суток наложила отпечаток и па процесс транспирации. Как устьичные движения, так и транспирация имеют свой определенный суточный ход. Все растения разделяются в отношении суточного хода устьичных движений на три группы: 1) растения, у которых ночью устьица всегда закрыты. Утром устьица открываются, и их дальнейшее поведение в течение дня зависит от условий среды. Мало воды — они закрываются. Достаточно воды — они открываются. К этой группе относятся в первую очередь хлебные злаки; 2) растения, у которых устьица утром открываются, а днем в зависимости от условий открыты или закрыты. Ночное их поведение зависит от дневного. Если днем устьица были закрыты, то ночью они открываются. Если днем они были открыты, то ночью они закрываются. К этой группе относятся растения с тонкими листьями — люцерна, горох, клевер; 3) растения, у которых ночью устьица всегда открыты (суккуленты), а днем, как и у всех остальных групп растений, открыты или закрыты в зависимости от условий.
Что касается суточного хода транспирации, то в ночной период суток транспирация резко сокращается. Это связано как с изменением внешних факторов (повышение влажности воздуха, снижение температуры, отсутствие света), так и с внутренними особенностями (закрытие устьиц). Измерения показывают, что ночная транспирация составляет всего 3—5% от дневной.
Дневной ход транспирации обычно следует за изменением напряженности основных метеорологических факторов (освещенности, температуры, влажности воздуха). Максимум транспирации будет приходиться на 12—13 ч. Это подчеркивает сравнительно малую зависимость транспирации от физиологического состояния растения. Ведущим в этом комплексе внешних воздействий будет напряженность солнечной инсоляции. Интересно, что растения с разным расположением листьев несколько различаются по суточному ходу транспирационного процесса. На листья, повернутые ребром к горизонту, солнечные лучи начинают падать раньше. В связи с этим подъем транспирации у таких растений в утренние часы также начинается несколько раньше.
В случае недостатка влаги кривая суточного хода транспирационного процесса из одновершинной превращается в двухвершинную. В полуденные часы транспирация сокращается, это позволяет растению восполнить недостаток воды, и тогда к вечеру транспирация снова возрастает.
При частом измерении транспирации можно заметить, что это пульсирующий процесс, т. е. ему свойственно ритмичное увеличение и уменьшение интенсивности. По-видимому, это связано главным образом с колебанием содержания воды в растении. Увеличение транспирации приводит к уменьшению содержания воды, что, в свою очередь, сокращает транспирацию. Как следствие, содержание воды растет и транспирация также возрастает, и так непрерывно [Козловский 2008-560].
Таким образом, на интенсивность испарения воды влияют условия, в которых находится растение: температура воздуха, освещенность, влажность почвы и воздуха, сила ветра. При высокой температуре, ярком солнце, небольшой влажности воздуха, сильном ветре испарение идет сильнее, чем в тихую пасмурную погоду.
Растения, обитающие в условиях недостатка влаги, выработали различные приспособления к ее излишней потере: утолщенные наружные стенки клеток кожицы листьев, восковой налет, густые волоски. Эти приспособления не только препятствуют испарению, но и способствуют отражению солнечных лучей. Кроме того, растения, постоянно испытывающие недостаток влаги, имеют небольшие листовые пластинки.
II Роль листьев в испарении воды растениями
2.1 Лист как орган транспирации
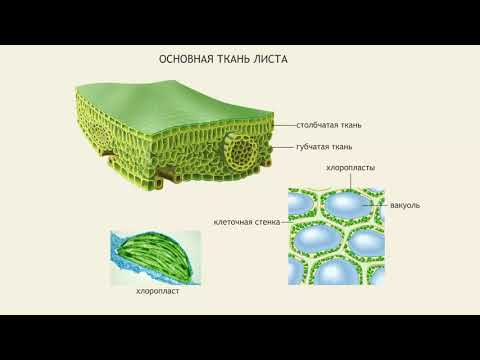
Основным транспирирующим органом является лист. Средняя толщина листа составляет 100-200 мкм. Паренхимные клетки листа расположены рыхло, между ними имеется система межклетников, составляющая в общей сложности от 15 до 25% объема листа. Эпидермис — покровная ткань листа, состоит из компактно расположенных клеток, наружные стенки которых утолщены. Кроме того, листья большинства растений покрыты кутикулой, в состав которой входят оксимонокарбоновые кислоты, содержащие по 16-28 атомов углерода и по 2-3 гидроксильные группы. Эти кислоты соединены друг с другом в цепочки с помощью эфирных связей. Кутикула варьирует как по составу, так и по толщине. Более развитой кутикулой характеризуются листья светолюбивых растений по сравнению с теневыносливыми и засухоустойчивых по сравнению с влаголюбивыми. Кутикула вместе с клетками эпидермиса образует как бы барьер на пути испарения паров воды. Особенно значительную преграду составляет кутикула. Удаление кутикулы во много раз повышает интенсивность испарения. Все эти особенности выработались в процессе эволюции как приспособление к сокращению испарения. Для соприкосновения листа с атмосферой имеются поры — устьица [ Красильникова 2009-347 ].
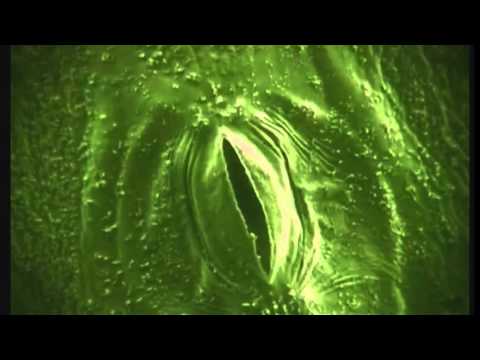
Устьице — это отверстие (щель), ограниченная двумя замыкающими клетками. Устьица встречаются у всех наземных органов растения, но больше всего у листьев. Каждая замыкающая клетка устьица имеет хлоропласты, в отличие от клеток эпидермиса. В них осуществляется фотосинтез, хотя с меньшей интенсивностью, чем в клетках мезофилла.
Устьица — одно из оригинальных приспособлений, обладающих способностью открываться и закрываться в зависимости от насыщенности замыкающих клеток водой. Обычно устьичные отверстия ограничены двумя замыкающими клетками, стенки которых неравномерно утолщены. У двудольных растений замыкающие клетки бобовидной, или полулунной, формы, при этом их внутренние прилегающие друг к другу стенки более толстые, а внешние — более тонкие. Когда воды мало, замыкающие клетки плотно прилегают друг к другу и устьичная щель закрыта. Когда воды в замыкающих клетках много, то она давит на стенки и более тонкие стенки растягиваются сильнее, а более толстые втягиваются внутрь, между замыкающими клетками появляется щель [Максимов 1978-670].
В последнее время показано, что для движения устьиц большое значение имеет также радиальное расположение микрофибрилл целлюлозы. Значение такого расположения было доказано на модели.
У злаков строение замыкающих клеток несколько иное. Они представлены двумя удлиненными клетками, на концах которых стенки более тонкие. При насыщении водой более тонкие стенки на концах растягиваются и раздвигают замыкающие клетки, благодаря чему образуется щель.
Число устьичных отверстий колеблется в зависимости от вида растений от 10 до 600 на 1мм 2 листа. У многих растений (75% видов), в том числе для большинства древесных, устьица расположены на нижней стороне листа. Диаметр устьичных щелей составляет всего 3-12мкм. Устьица соединяют внутренние пространства листа с внешней средой. Вода поступает в лист через сеть жилок, в которых расположены сосудистые элементы. Возможны три пути испарения: через устьица — устьичная, через кутикулу — кутикулярная и через чечевички — лентикулярная транспирация. Впервые разграничение на кутикулярную и устьичную транспирацию было введено в 1877г.
В том, что действительно испарение идет не только через устьица, но и через кутикулу, легко убедиться. Так, если взять листья, у которых устьица расположены только с нижней стороны (например, листья яблони), и замазать эту сторону вазелином, то испарение воды будет продолжаться, хотя и в значительно уменьшенном размере. Следовательно, определенное количество воды испаряется через кутикулу. Кутикулярная транспирация обычно составляет около 10% от общей потери воды листом. Однако в некоторых случаях у растений, листья которых характеризуются слабым развитием кутикулы, доля этого вида транспирации может повышаться до 30%. Имеет значение также возраст листа. Молодые листья, как правило, имеют слабо развитую кутикулу и, следовательно, более интенсивную кутикулярную транспирацию. У старых листьев доля кутикулярной транспирации снова возрастает, так как, хотя кутикула и сохраняет достаточную толщину, в ней появляются трещины, через которые легко проходят пары воды.
Трещины в кутикуле могут появляться и после временного завядания листьев, благодаря чему транспирация усиливается. Имеются данные, что кутикулярная транспирация меньше зависит от условий внешней среды по сравнению с устьичной (Н. И. Антипов). Основная часть воды испаряется через устьица. Процесс транспирации можно подразделить на ряд этапов. Первый этап — это переход воды из клеточных оболочек, где она находится в капельножидком состоянии, в межклетники (парообразное состояние). Это собственно процесс испарения, отрыв молекул воды с поверхности клеточных стенок. Важно подчеркнуть, что уже на этом этапе растение обладает способностью регулировать процесс транспирации (внеустьичная регулировка). Так, если в растении недостаток воды, то в сосудах корня и стебля создается сильное натяжение, которое делает их водный потенциал более отрицательным, что оказывает сопротивление передвижению воды в клетку и уменьшает интенсивность испарения. Надо учитывать также, что между всеми частями клетки существует водное равновесие. Чем меньше воды в клетке, тем выше становится концентрация клеточного сока. А это, в свою очередь, уменьшает содержание свободной воды в протопласте и клеточной оболочке. Соотношение свободной воды к связанной падает, водоудерживающая сила растет, интенсивность испарения уменьшается. Второй этап — это выход паров воды из межклетников или через кутикулу, или, главным образом, через устьичные щели. Поверхность всех клеточных стенок, соприкасающихся с межклетными пространствами, превышает поверхность листа примерно в 10-30 раз. Все же если устьица закрыты, то все это пространство быстро насыщается парами воды и переход воды из жидкого в парообразное состояние прекращается. Иная картина наблюдается при открытых устьицах. Как только часть паров воды выйдет из межклетников через устьичные щели, так сейчас же этот недостаток восполняется за счет испарения воды с поверхности клеток. Поскольку устьичная транспирация составляет 80-90% от всего испарения листа, то степень открытости устьиц является основным механизмом, регулирующим интенсивность транспирации. При открытых устьицах общая поверхность устьичных щелей составляет всего 1-2% от площади листа. Казалось бы, это должно очень сильно уменьшать испарение по сравнению с испарением свободной водной поверхности той же площади, что и лист. Однако это не так. Сравнение испарения листа с испарением со свободной водной поверхности той же площади показало, что оно идет не в 100 раз, как эта следовало, бы, исходя из размеров открытой площади (1%), а всего в 2 раза медленнее. Объяснение этому явлению было дано в исследованиях английских ученых Броуна и Эскомба, которые установили, что испарение из ряда мелких отверстий идет быстрее, чем из одного крупного той же площади. Это связано с явлением краевой диффузии. При диффузии из отверстий, отстоящих друг от друга на некотором расстоянии, молекулы воды, расположенные по краям, рассеиваются быстрее. Естественно, что таких краевых молекул значительно больше в ряде мелких отверстий по сравнению с одним крупным.
Для малых отверстий интенсивность испарения пропорциональна их диаметру, а не площади. Указанная закономерность проявляется в том случае, если мелкие поры расположены достаточно далеко друг от друга. Структура листа удовлетворяет указанным требованиям. Поры (устьица) имеют малый диаметр и достаточно удалены друг от друга. При открытых устьицах выход паров воды идет достаточно интенсивно, закрытие устьиц резко тормозит испарение. Именно на этом этапе вступает в действие устьичная регулировка транспирации. При недостатке воды в листе устьица автоматически закрываются. Полное закрывание устьиц сокращает транспирацию примерно на 90%. Вместе с тем уменьшение диаметра устьичных щелей не всегда приводит к соответственному сокращению транспирационного процесса. Определения показали, что устьица должны закрыться больше чем на 1/2, для того чтобы это сказалось на уменьшении интенсивности транспирации.
Третий этап транспирации — это диффузия паров воды от поверхности листа в более далекие слои атмосферы. Этот этап регулируется лишь условиями внешней среды [ Радкевич 2005-235 ].
2.2 Биологическое значение транспирации
Количество воды, испаряемой растением, во много раз превосходит объем содержащейся в нем воды. Экономный расход воды составляет одну из важнейших проблем сельскохозяйственной практики. К. А. Тимирязев назвал транспирацию, в том объеме, в каком она идет, необходимым физиологическим злом. Действительно, в обычно протекающих размерах транспирация не является необходимой. Так, если выращивать растения в условиях высокой и низкой влажности воздуха, то, естественно, в первом случае транспирация будет идти со значительно меньшей интенсивностью. Однако рост растений будет одинаков или даже лучше там, где влажность воздуха выше, а транспирация меньше. Вместе с тем транспирация в определенном объеме полезна растительному организму:
1. Транспирация спасает растение от перегрева, который ему грозит на прямом солнечном свете. Температура сильно транспирирующего листа может примерно на 7°С быть ниже температуры листа завядающего, нетранспирирующего. Это особенно важно в связи с тем, что перегрев, разрушая хлоропласты, резко снижает процесс фотосинтеза (оптимальная температура для процесса фотосинтеза около 30-33°С). Именно благодаря высокой транспирирующей способности многие растения хорошо переносят повышенную температуру.
2. Транспирация создает непрерывный ток воды из корневой системы к листьям, который связывает все органы растения в единое целое.
3. С транспирационным током передвигаются растворимые минеральные и частично органические питательные вещества, при этом, чем интенсивнее транспирация, тем быстрее идет этот процесс.
Как уже говорилось, механизм поступления питательных веществ и воды в клетку различен. Однако некоторое количество питательных веществ может поступать пассивно, и этот процесс может ускоряться с увеличением транспирации [Якушина 2008-280].
III Практическая работа на тему «Определение общей площади поверхности листьев дуба, березы и тополя»
«Определение общей площади поверхности листьев дуба, березы и тополя» [Жданов 2006-450].
При выполнении практической работы была использована следующая инструкция:
1. Возьмем лист березы с обратной стороны в клеточку. Примем одну клеточку за 1 см.
2. Подсчитаем число полных квадратов (а), попавших внутрь контура. Запишем в таблицу показания.
3. Подсчитаем число неполных квадратов ( b ), пересеченных контуром. Запишем показания в таблицу.
4. Разделим число неполных квадратов пополам. Внесем показания в таблицу.
5. Найдем общую площадь листовой поверхности по формуле: S = a + b /2, где S -площадь, — сумма.
6. Сделаем тоже самое с листом дуба и тополя. [Приложение №2].
1.Самая большая площадь поверхности у листа дуба, поэтому с этого листа испарение идет интенсивнее.
2. Самая маленькая площадь у листа березы, поэтому с этого листа процесс испарения менее интенсивный.
3. Лист тополя занимает промежуточную позицию, между листом дуба и листом березы.
В данном реферате рассмотрены особенности одного из важных процессов наряду с фотосинтезом, дыханием – это транспирация, то есть испарение воды растениями. Процесс транспирации представляет собой движение воды через растения, и ее испарение через наружные органы, такие как листья, стебли и цветы. К условиям, влияющим на испарения относятся: температура воздуха, освещенность, влажность почвы.
Биологическая значимость транспирации заключается в том, что транспирация спасает растения от перегрева, создает непрерывный ток воды из корневой системы к листьям, с транспирационным током передвигаются растворимые минеральные вещества, при этом, чем интенсивнее транспирация, тем быстрее идет этот процесс. Все это опровергает точку зрения, что рост растений будет одинаков или даже лучше там, где транспирация меньше. Однако могут быть исключения, а именно растение как кактус может расти в пустыне без влаги.
Следовательно, транспирация – жизненно-важный процесс для растений. Он способствует увеличению продолжительности жизни растений.
Доказано, что большая поверхность листьев имеет огромное значение для питания растений и для испарения большого количества воды. Проведя практическую работу, была выявлена площадь поверхности трех листьев дуба, березы и тополя. Таким образом, можно определить уровень интенсивности испарения у листьев разных растений и сделать сравнительный анализ, у какого растения процесс испарения быстрее, а у какого медленнее. У дуба процесс испарения гораздо выше чем у березы и тополя, что защищает это растение от перегрева и высыхания. Вот почему, дуб можно увидеть в жарких странах, например, в Тунисе. Береза и тополь встречается в менее жарких странах, например, они широко распространены в России, так как их уровень транспирации невысокий и в жарких странах они могут засохнуть.
Следовательно, рост растений будет лучше там, где транспирация выше. Уровень транспирации зависит от площади листа. Листья с большой поверхностью наблюдаются в жарких странах, так как благодаря большой поверхности листа ускоряется процесс транспирации и растение не засыхает. В менее засушливых районах площадь поверхности листа не имеет значения, так как при высокой влажности воздуха уходит потребность в большой поверхности листа. Поэтому площадь таких листьев как береза и тополь мала. Но когда наступает засушливое лето. Листья не справляются с транспирацией и они засыхают и можно увидеть ранний листопад.
1. Геннель П.А. Физиология устойчивости растительных организмов./П.А.Геннель. – М.:МГУ,2010-470с.
2. Горышена Т.К. Экология растений./Т.К.Горышена. – М.:Высшая школа,2011 – 345с.
3. Гэлстон А.П. Жизнь зеленого растения./А.П. Гэлстон. – М.: Мир, 2009 – 380с.
4. Жданов В.С. Испарение воды листьями./В.С. Жданов. – М.: МГУ, 2006 – 450с.
5.Козловский П.К. Водный обмен растений./П.К.Козловский. – М.: Колос, 2008 – 560с.
6. Красильникова Н.А. Жизнь растений./Н.А. Красильникова.-М.: Просвещение, 2009 -347с.
7. Максимов Н.А. Водный режим растений./Н.А. Водный режим растений. – М.: Советская энциклопедия, 1978 -670с.
8. Радкевич В.А. Экология./В.А. Радкевич. – Минск.: Высшая школа,2005 – 235с.
9.Якушина Н.И. Физиология растений./Н.И. Якушина. – М.: Просвещение,2008 – 280с.
💡 Видео
Биология. 9 класс. Внешние и внутренние факторы, влияющие на транспирацию /27.10.2020/Скачать

Площадь поверхности призмы. Практическая часть. 11 класс.Скачать

Строение листаСкачать
Лист. Обнаружение дыхания листьевСкачать

Схема работы устьиц листаСкачать

Биология. Урок №14, Лаб раб №4Скачать

Органы дыхания у растений УстьицаСкачать
Определение интенсивности транспирации и относительной транспирации весовым методомСкачать

Биология. 10 класс. Влияние соотношения площади поверхности к объему на скорость диффузииСкачать

Испарение воды листьямиСкачать

Влияние среды на строение листа. Видоизменение листьев | Биология 6 класс #27 | ИнфоурокСкачать

Биология. урок №15Скачать

Влияние внешних факторов на транспорт веществ по флоэме. 9 класс.Скачать

Анатомия растений. Парадермальный срез листа и изготовление реплики эпидермиса (А. Платонова)Скачать

Испарение воды листьями. Листопад. Видеоурок по биологии 6 классСкачать

6 класс Биология - Испарение воды растениями (транспирация). Листопад.Скачать

ТранспирацияСкачать
